Abstract
Previous studies by Stachel and colleagues indicate that lithium chloride induction of post-midblastular Brachydanio rerio embryos results in deficiencies in normal anterior-posterior development. To determine whether or not higher concentrations of lithium chloride lead to successively greater defects in anterior-posterior development, D. rerio embryos were induced with the following variable concentrations of lithium chloride: 0.45 M, 0.30 M, 0.15 M, and 0.00 M. Embryos induced with 0.15M of LiCl exhibited little or no phenotypic deviation from the control group, whereas 0.30 M and 0.45 M LiCl-induced embryos did not develop eyes and showed signs of perturbed tail development.The results of this experiment suggest that the degree of inhibition of normal anterior-posterior development increased in embryos induced with higher concentrations of lithium chloride. Lithium chloride induction may be responsible for the reduced transcription of goosecoid in the anterior regions of D. rerio or the prevention of cell movements allowing the anterolateral accumulation of goosecoid, resulting in the perturbation of normal anterior development (Stachel et. al., 1993).
Introduction
Lithium chloride is a known teratogen which alters development in a variety of organisms including sea urchins, Xenopus (frogs), andBrachydanio rerio (Zebrafish) (Gilbert, 2003). In sea urchin embryos, lithium chloride causes the accumulation of nuclear Beta-catenin in every cell, and transforms presumptive ectoderm into endoderm (Gilbert, 2003). Lithium exposure in cleavage-stage embryos ofXenopus inhibits dorsal/ventral axis specification and results in radially-symmetric, dorsal-anteriorized embryos (Stachel et. al., 2003). The research conducted by Stachel and colleagues suggests that lithium induction of pre-midblastular Zebrafish prevents normal dorsal/ventral axis patterning by acting as an inhibitor to the phosphoinositol pathway, which results in goosecoid and nogginexpression outside the region of the presumptive embryonic shield instead of these genes being confined to the region proximal to the dorsal blastopore lip.
Experiments by Stachel and colleagues have shown that the development of anterior structures is dependent on Wnt signaling, especially the transcription of goosecoid, which codes for a dorsalizing protein necessary for normal anterior development (Stachel et. al., 2003). The increase in gene expression of organizer-specific proteins such as Goosecoid in the presumptive ventral regions of the organism produces different phenotypic results when the induction occurs at certain stages in development (Stachel et. al., 1993). For instance, the exposure of embryos to LiCl before the midblastular transition (2 hour stage) results in hyperdorsalization and the inhibition of normal dorsal/ventral axis patterning (Deitrich, 1999). In contrast, embryos exposed to LiCl at the four-hour stage after the midblastular transition experienced normal dorsal/ventral axis specification but perturbed development of anterior structures such as eyes (Stachel et. al., 1993).
The purpose of this experiment is to study the effects of lithium teratogenesis on post-midblastular embryo to observe whether or not the inhibition of anterior development occurs along a gradient, with increasing concentrations of lithium chloride leading to incrementally more severe defects in anterior development. Alternatively, there may be a certain threshold at which lithium induces defects in anterior development. Indications of the former hypothesis would consist of lithium teratogenesis leading to, for example, partial formation of an eye at one concentration and the complete absence of eye formation at a slightly higher concentration, then it may be possible that lithium teratogenesis affects development along a gradient. In accordance with the latter situation, the observable effects of lithium teratogenesis would consist in the complete absence of the eyes at one concentration and complete formation and presence of all eye structures including retina and lens at a slightly lower concentration.
Objective
The purpose of this experiment is to monitor the effects of different concentrations of LiCl on anterior development in zebrafish embryos. This will be carried out by assessing the degree to which certain amounts of the lithium cholride teratogen affect the morphology of anterior structures such as the eye.
Materials
60-80 Brachydanio rerio embryos at the sphere/dome-stage of development (see procedure for more details)
LiCl (0.15 M, 0.30 M, and 0.45 M) in Zebrafish embryo medium
distilled H2O
9 60 mm glass petri dishes
wide-mouth pastuer pipette (with large enough opening for suctioning chorionated zebrafish eggs)
dissection scope
Procedure
1. Obtain 15-20 zebrafish embryos at the sphere/dome-stage of development (four hours post-fertilization) for each of the four titrations of lithium chloride to be tested, inclusive of the control group. Therefore, about 60-80 sphere/dome-stage embryos must be isolated in total. For a description of sphere/dome-stage embryos, refer to the Zebrafish Information Network homepage (www.zfin.org).
2. Place the embryos into four separate petri dishes, each containing 10 mL of Zebrafish embryo medium.
3. Next, procure three dishes and fill them with the following concentrations of LiCl:
0.45 M
0.30 M
0.15 M
A table including the calculations for the three LiCl titrations used in the original experiment is included in the Materials and Methods section of this page.
4. Transfer each population of embryos from the embryo medium to the LiCl-containing dishes using a severed pipette and immerse them in solution for 10 minutes.
5. After 10 minutes, rinse the embryos by placing each group in a separate dish containing 10 milliliters of embryo medium, and then transfer back to the original embryo-medium-containing dishes. Be sure to label the dishes with the corresponding amount of LiCl to which they were induced.
6. Photograph the embryos after 24 hours to observe anterior development, carefully noting deviations in anterior patterning and formation from the control population.
Materials and Methods
The embryos used in this experiment were at the sphere stage of development, which is equivalent to approximately four hours of incubation post-fertilization. More importantly, the sphere stage occurs after the midblastular transition, whereupon the embryo begins to translate its own genes, cell movements occur, and a pattern of slow, asynchronous cleavage begins (Gilbert, 2003). The exposure of each of the variant groups to lithium chloride consisted of a ten-minute incubation period for each so that the time of incubation would remain concurrent with the sphere stage and not extend beyond this period of development. Sphere-stage embryos were isolated according to Stages of Embryonic Development of the Zebrafish (Zebrafish Information Network, 1995). Isolated embryos were collected in Zebrafish Embryo Medium, which was prepared according to the developmental biology lab website (DB Lab, 2003). During the induction perion, approximately 10-15 Brachidanio rerio embryos were treated with the following titrations of LiCl: 0.45 M, 0.30 M, and 0.15 M made up in Zebrafish Embryo Medium. The calculations for the concentrations of Lithium Chloride that were used are as indicated in Table I:
Table I: Variable Molarity of Lithium Chloride Titrations
The fourth group did not contain any LiCl and served as a control, thus no calculations were required for this titration. Subsequent to the ten-minute incubation period of the three variable groups in the LiCl solutions, all embryos were rinsed with Zebrafish embryo medium and incubated overnight at 28°C.
Results
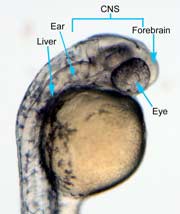
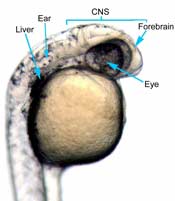
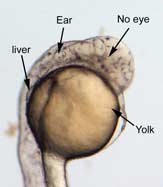
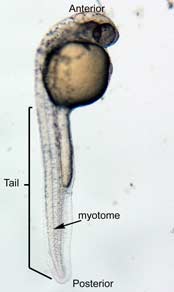
In the control group embryos, anterior-posterior patterning was normal, with full formation of the terminus structures, the retina, and the lens of the eye (Figure 1A). In embryos treated with 0.15 M LiCl, the eyes appear to have developed normally, but the eye shown in Figure 1B is relatively closer to the anterior terminus than the eye in Figure 1A. The 0.15 M LiCl-treated embryos exibited slightly stunted growth at their anterior and posterior termini; the tails were also slightly bent and rounded at the tip (Figure 2B. The 0.30 M LiCl-treated embryos did not develop eyes, and exhibited slight curvature of the dorsal region (Figure 1C). These embryos exhibited even less development of the anterior and posterior termini than the 0.15 M embryos, with stubby tails and large yolk sacs (Figure 2C).
All embryos in Figure1 exhibit development of ears, liver, and the CNS. The average survival rate for embryos in the 0.00 M, 0.15 M, and 0.30 M solutions were 19/19, 18/20, and 15/21 respectively. The 0.45 M LiCl -treated embryos had lower viability, with only seven of the original nineteen embryos surviving the 24-hour period from the time of incubation to the time of data collection. The resulting embryos were fragile, and some did not survive the dechorionation process and their cells became disaggregated by the pull of the forceps and the influx of Zebrafish embryo medium into the opened chorion. This group displayed a lack of anterior-posterior development, with no definable head structures, and stunted tail development (Figure 2D).
|
|
|
A 0.00 M LiCl | B 0.15 M LiCl | C 0.30 M LiCl |
Figure 1 (A) Control embryo, typical anterior/posterior patterning with fully-formed anterior structures including the eye, forebrain, CNS, ear and liver (B) Embryo induced with 0.15 M LiCl shows liver, ear, eye, forebrain, and CNS development (C) 0.30 M LiCl-induced embryo shows no eye formation, though ear and liver are present.
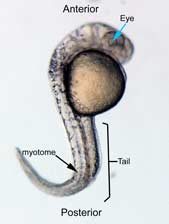 |  |
A 0.00 M LiCl | B 0.15 M LiCl |
C 0.30 M LiCl D 0.45 M LiCl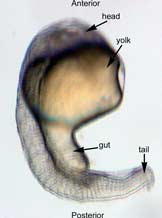
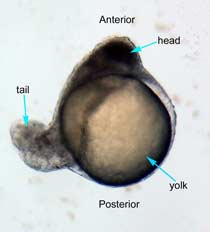
Figure 2 Photographs taken 26 hours after LiCl induction (A) Control embryo shows normal tail formation (B) Embryo shows normal tail development (C) Tail appears stunted and crooked, gut is shortened, head is smaller, and yolk is enlarged and misshapen (D) Head and tail are stunted, anterior structures are missing, yolk is enlarged.
Discussion
These results demonstrate that lithium chloride treatment affected the anterior-posterior morphogenesis of Brachydanio rerio embryos such that higher concentrations of this substance lead to increasingly greater inhibition of anterior and posterior development. The 0.15 M LiCl-treated embryo were missing their anteriormost structures. The embryos treated with 0.30 M LiCl exhibited greater teratogenic effects on the development of anterior and posterior terminus structures. Higher concentrations of LiCl also led to more morphological defects in both the anterior and posterior terminal regions, further confirming the hypothesis that the concentrations of LiCl teratogen is directly related to the phenotypic effect on Zebrafish.
Lithium-induced teratogenesis has been shown to perturb the development of ventral structures in premidblastular Zebrafish, when the blastula is still forming morphogenetic gradients of proteins in the Wnt signaling pathway (such as b-catenin) and continuing to transcribe maternal mRNAs. In these early embryos, lithium teratogenesis leads to the expression of bustled and radialized phenotypes, which exhibit hyperdorsalization (Stachel et. al., 1993). In the sphere-stage embryos, morphogenetic gradients have already been established, so the effect of lithium in this case probably occurs downstream of these events.
Studies conducted by Stewart and Gerhart (1990) showed that lithium inhibits gastrulation m
日前,國產期刊TheInnovation獲得首個影響因子(IF=32.1),成為科睿唯安JCR綜合性期刊分類下排名僅次于《自然》(IF=64.8)和《科學》(IF=56.9)的期刊,并且這本期刊在目前......
近日,服務科學領域的全球領導者賽默飛世爾科技(以下簡稱賽默飛)宣布,在達成收購意向兩個月之后,賽默飛以28億美元、折合人民幣約190億元的價格,完成了對TheBindingSiteGroup的全現金收......
11月15日,施普林格·自然和TheLens平臺宣布結成重要的合作伙伴關系,以更深入地揭示學術研究和數據如何能通過經濟和社會成效,加速推動創新的問題解決方式。通過將科學、投資和企業領域的開放數據更好地......
蛋白質作為構成人體組織器官的支架和主要物質,在人體生命活動中起著重要作用。蛋白質的相互作用能產生許多效應,如形成特異底物作用通道、生成新的結合位點、失活、作用底物專一性和動力學變化等,細胞的代謝、信號......
2021年9月9日,無錫臻和生物科技有限公司(以下簡稱“臻和科技”)與美國VyantBio公司簽署TissueofOrigin?(以下簡稱“TOO?”)全球權益和ZL轉讓協議,全資收購這款唯一獲FDA......
2021年7月20日,JournalofCellularPhysiology及JournalofCellularBiochemistry同時撤回了中國學者49篇文章。從2019年開始,Journalo......
6月10日,QS教育集團正式發布了2021年世界大學排名,中國共有83所高校上榜,包括內地高校51所,港澳臺地區高校32所。中國大學的總體排名情況已經連續數年呈上升趨勢,今年再度刷新了榜單。大學排名,......
磷酸甘油酸突變酶1(PGAM1)通過其代謝活性以及與其他蛋白質(例如α平滑肌肌動蛋白(ACTA2))的相互作用,在癌癥代謝和腫瘤進展中起關鍵作用。變構調節被認為是發現針對PGAM1的高選擇性和有效抑制......
作為一種重要的植物激素,茉莉酸不僅調控植物對于機械損傷、昆蟲取食和腐生型病原菌侵害的防御反應,還參與調控諸多生長發育過程。basicHelix-Loop-Helix(bHLH)類型轉錄因子MYC2是茉......
ThePlantCell是植物領域的著名學術期刊,對植物學的發展起到了重要的引領作用。為慶祝創刊30周年,ThePlantCell雜志社邀請部分編委會成員及其他科學家對發表在該雜志的重要研究工作進行評......