經典的生物研究技術側重于分子和細胞集群的研究——即研究含有大量相同形態或功能的分子或細胞的活動。但是,這種方法會忽略集群中的單個分子或子群的特異性。事實上在細胞周期的不同階段或在不同的環境中,單個分子或細胞的活動很可能與集群表現出的整體活動不同。要對單個分子或亞群的活動進行觀察,必須嚴格控制實驗條件,保證每個分子的狀態相同。
單分子熒光成像技術可以分為兩類:一類是在外力作用下研究單分子活動,通常通過原子力顯微鏡(AFM)、光鑷(OT)或磁鑷(MT)將力施加到單個分子上。另一類就是用熒光顯微成像觀察生物系統中單分子活動。
熒光顯微成像是生命科學領域觀察生物體結構的經典方法。這其中,用熒光探針標記,檢測和分析單個分子的單分子熒光成像技術,能夠幫助科學家們在不破壞生命體正常生理狀態的情況下,清晰地觀察到單個分子的活動。

我們常用的寬場顯微鏡能夠同時觀察數百個分子和它們之間的相互作用,使研究人員能夠輕松地收集到大量數據進行分析,同時還增加了檢測到小概率事件的可能性。但是由于它不能排除來自焦平面上下的雜散信號,信噪比較差(圖1),對于本身信號已經很弱的單分子熒光來說,寬場成像不是一種好的成像方法。

圖1 普通熒光和TIRF照明下表達GFP的Hela細胞在同一焦平面的圖像。Scale bar: 10 μm
共聚焦顯微鏡可以克服寬場顯微鏡的問題,拒絕雜散光,這樣就可以只對感興趣的平面成像。由于共聚焦顯微技術的特點,科學家們將它應用于DNA修復等領域,直接觀察核苷酸切除修復蛋白與DNA的結合(Segers-Nolten et al. 2002)。
TIRF
全內反射熒光顯微鏡(TIRF)是單分子熒光成像最常用的方法之一,被用于研究肌動蛋白和肌球蛋白轉運動力學(Zimmerman et al.)以及分子擴散(Fujiwara et al.,2016)等。簡單來說,TIRF是利用光線全反射后在介質另一面產生衰逝波的特性,使用特定角度的激發光,令所有的光都被反射。這樣在全反射區域的另一面就會產生衰逝波,對樣本表面的極薄區域(通常在200nm以下)進行照明(圖2)。由于衰逝波是呈指數衰減的,只有極靠近全反射面的樣本區域會被激發,從而大大降低了背景噪聲,提高了信噪比(圖1)。TIRF廣泛應用于對細胞表面物質的動態觀察,如固定在蓋玻片或細胞膜表面上的分子等。

圖2 傳導波和衰逝波的差別。傳導波(propagating wave)以線性方式在組織中傳播,可激發深度大于1 mm的熒光基團。由TIRF產生的衰逝波(Evanescent wave),激發深度僅為約100nm
FRET
如果一個熒光分子(供體分子)的發射光譜與另一個熒光分子(受體分子) 的吸收光譜有一定的重疊, 當這兩個分子的距離足夠近(通常為1-10nm)的時候,供體的熒光能量可以向受體轉移,表現為受體的熒光增強,同時供體自身的熒光衰減。這就是熒光共振能量轉移(FRET)。FRET 程度與供、受體分子的空間距離緊密相關,隨著距離延長, FRET顯著減弱。而FRET速率與兩個分子間的距離成正比。利用這種供體和受體分子之間非輻射能量轉移的原理,FRET可以作為“光譜標尺”,用來確定兩個熒光分子的接近程度。
Theodor F?rster在1960年最早提出了FRET,并通過Stryer & Haugland(1967)等人的實驗得到證實。通過單分子FRET技術,我們能在單個分子內或分子復合物中,以納米尺度實時追蹤變化,了解目標分子在細胞內的空間分布,從而幫助解答許多重要的生物學問題。目前單分子FRET已經成為研究蛋白質&核酸結構和動力學的理想技術。包括觀察酶在其功能周期中的構象變化 (Dyla et al. 2016),實時監測DNA合成 (Fijen et al. 2017)等。
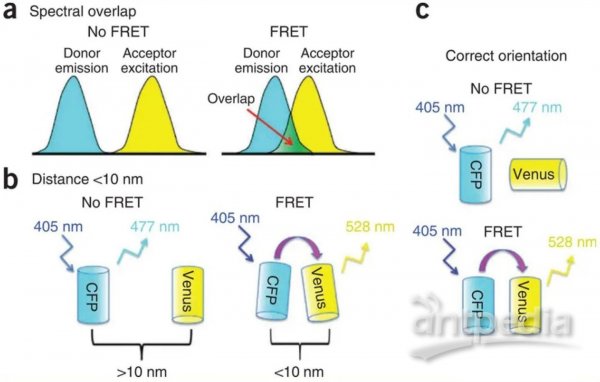
圖3 發生FRET必須滿足的三個最重要的條件:(a)供體的發射光譜必須與受體的激發光譜重疊。(b)供體和受體距離<10nm(c)供體和受體熒光雙極相互平行。(Broussard et al.2013年)
PALM/STORM
除了TIRF和FRET,我們在超分辨專題中給大家介紹的光激活定位顯微技術(PALM)和隨機光學重建顯微技術(STORM)也是單分子熒光成像技術。這里就不多做贅述,大家可以閱讀我們之前的技術文章前沿顯微成像技術專題——超分辨顯微成像(1)復習一下~
由于單個熒光基團的信號強度本身已經很弱,對信噪比的挑戰很大。有時單分子成像還需要很快的速度,曝光時間較短,進一步降低信號強度。因此,決定單分子熒光成像質量的一個最重要的因素就是能否從背景噪聲中檢測出微弱的信號——這就需要高靈敏度、低噪聲和高速的檢測設備了。
EMCCD 曾經是單分子熒光成像的首選,這是因為它95%的量子效率和電子倍增過程帶來的高信噪比。然而,由于存在增益衰減,EMCCD 的成像效果不穩定,信噪比也會收到電子倍增過程帶來的額外噪聲的影響而降低。
隨著科學相機技術的飛速發展,背照式 sCMOS 相機誕生了。它和 EMCCD 一樣具有95%量子效率和極低的讀出噪聲,但是不存在EMCCD增益衰減和額外噪聲的問題,因此能夠保證以極高的信噪比檢測單分子熒光信號。同時,背照式 sCMOS在速度、視野和性價比上都具有很大的優勢,因此,近年來越來越多的單分子熒光成像應用已經“移情別戀”,使用背照式 sCMOS 取代傳統的 EMCCD,取得了非常好的成像效果。
單分子熒光技術的基礎是用熒光染料對感興趣的分子進行標記,標記方法和熒光染料的選擇至關重要,需要根據樣品和特定成像技術的需要進行選擇。標記單個分子的方法有很多,包括抗體標記、生物素化、表位標記、小分子探針和生物正交標記等。一般來說,用于單分子成像的熒光染料應該滿足以下要求:
熒光效率高,實現信號最大化
熒光穩定(不快速光漂白),允許較長時間成像
熒光分子小,不破壞目標分子的生物活性
發射光譜在可見光區域,最好是在相機量子效率峰值的區域。
常見的熒光染料包括有機染料(如FITC、TRITC)、熒光蛋白(如GFP、YFP)和量子點。當然,這些染料并不是完美的,也有著各自的優缺點,我們將在后面的文章中給大家做詳細的介紹。
與集群研究相比,單分子熒光成像能夠揭示單個分子的結構和功能。自從20世紀90年代單分子成像技術被廣泛應用以來,已經開發出許多新技術,包括效率更高的熒光染料和更靈敏的科學相機,使得單分子實驗中信噪比低的問題得到了很好的解決。這也使得單分子熒光成像越來越受到歡迎。
REFERENCES
· Ishikawa-Ankerhold, H. C., Ankerhold, R. & Drummen, G. P. (2012) Advanced fluorescence microscopy techniques--FRAP, FLIP, FLAP, FRET and FLIM. Molecules. Apr 2;17(4):4047-132. doi: 10.3390/molecules17044047
· Shashkova, S. & Leake, M. C. (2017) Single-molecule fluorescence microscopy review: shedding new light on old problems. Biosci Rep. Jul 21;37(4). pii: BSR20170031. doi: 10.1042/BSR20170031
· Broussard, J. A., Rappaz, B., Webb, D. J. & Brown, C. M. (2013) Fluorescence resonance energy transfer microscopy as demonstrated by measuring the activation of the serine/threonine kinase Akt. Nat Protoc. Feb;8(2):265-81. doi: 10.1038/nprot.2012.147
· Dyla, M., Andersen, J. L., Kjaergaard, M., Birkedal, V., Terry, D. S., Altman, R. B., Blanchard, S. C., Nissen, P. & Knudsen, C. R. (2016) Engineering a Prototypic P-type ATPase Listeria monocytogenes Ca(2+)-ATPase 1 for Single-Molecule FRET Studies. Bioconjug Chem. Sep 21;27(9):2176-87. doi: 10.1021/acs.bioconjchem.6b00387
· Fijen, C., Montón Silva, A., Hochkoeppler, A. & Hohlbein, J. (2017) A single-molecule FRET sensor for monitoring DNA synthesis in real time. Phys Chem Chem Phys. Feb 8;19(6):4222-4230. doi: 10.1039/c6cp05919h.
· Fujiwara, T. K., Iwasawa, K., Kalay, Z., Tsunoyama, T. A., Watanabe, Y., Umemura, Y. M., Murakoshi, H., Suzuki, K. G., Nemoto, Y. L., Morone, N. & Kusumi, A. (2016) Confined diffusion of transmembrane proteins and lipids induced by the same actin meshwork lining the plasma membrane. Mol Biol Cell. Apr 1;27(7):1101-19. doi: 10.1091/mbc.E15-04-0186
· Segers-Nolten, G. M., Wyman, C., Wijgers, N., Vermeulen, W., Lenferink, A. T., Hoeijmakers, J. H., Greve, J. & Otto, C. (2002) Scanning confocal fluorescence microscopy for single molecule analysis of nucleotide excision repair complexes. Nucleic Acids Res. Nov 1;30(21):4720-7.
· Stryer, L. & Haugland, R. P. (1967) Energy transfer: a spectroscopic ruler. Proc Natl Acad Sci U S A. Aug; 58(2): 719–726.
· Zimmermann, D., Santos, A., Kovar, D. R., Rock, R. S. (2015) Actin age orchestrates myosin-5 and myosin-6 run lengths. Curr Biol. Aug 3;25(15):2057-62. doi: 10.1016/j.cub.2015.06.033.
公告內容為便于供應商及時了解政府采購信息,根據《財政部關于開展政府采購意向公開工作的通知》(財庫〔2020〕10號)等有關規定,現將本單位2025年10月至2025年11月采購意向公開如下:序號采購項......
......
——徠卡中高端顯微鏡本土化交付慶典訪談2025年8月29日,上海浦東金橋工業區的徠卡金橋工廠洋溢著熱烈的氛圍。徠卡顯微系統(上海)貿易有限公司(以下簡稱“徠卡”)隆重舉辦“創升中國顯微新境”——DM4......
●蔡司蘇州研發制造基地開業一周年,持續推動本土創新和智能制造,現已成為蔡司全球顯微鏡業務的戰略樞紐。● 三款高端顯微鏡產品——蔡司激光共聚焦顯微鏡LSM910,蔡司場發射掃描電子顯微鏡Sig......
2025年8月29日,徠卡顯微系統(上海)貿易有限公司(以下簡稱“徠卡”)在上海金橋工廠舉行中高端顯微鏡本土化交付慶典,首批國產DM4/6/i8顯微鏡正式下線。近百位專家、學者及政府領導齊聚一堂,共同......
近日,由奧譜天成作為主要起草單位的兩項團體標準《植物表型葉綠素熒光成像分析設備》(我司排名第2)《基于無人機高光譜遙感的植物監測技術要求》(我司排名第3)正式發布,為生態遙感應用領域提供了系統性、權威......
zycgr210713012025年8至12月政府采購意向為便于供應商及時了解政府采購信息,根據《財政部關于開展政府采購意向公開工作的通知》(財庫〔2020〕10號)等有關規定,現將zycgr2107......
浙江豪圣建設項目管理有限公司受西湖大學的委托,就所需的冷凍雙束顯微鏡等5套設備(臨床醫學研究中心超長期國債項目)進行公開招標,歡迎符合條件的單位前來投標。具體如下:一、項目編號:WLU-HW-GK-W......
重慶電子科技職業大學集成電路與電子元器件失效檢測公共服務中心設備采購(一期)項目截止時間2025-08-1218:00采購項目概況本項目擬建設集成電路成品級和晶圓級的測試、集成電路失效分析實驗室,按照......
近期,為深入實施創新驅動發展戰略,加快建設具有全球影響力的科技創新中心,上海市科學技術委員會發布《2025年度關鍵技術研發計劃“科學儀器”項目申報指南》(以下簡稱《指南》)。根據《指南》要求,此次申報......